
植物遺伝育種学研究室での植物生殖機構の分子遺伝学的研究
1. 自家不和合性の分子遺伝学的研究
アブラナは自家不和合性という興味深い性質を持っています。これは、自分の花粉で受粉(自家受粉)すると種子が出来ないが、別の個体の花粉で受粉(他家受粉)すると種子が出来るという性質で、近親交配を防ぐために植物が持っている機構です(Kitashiba and Nasrallah 2014 Breed Sci 64, 23-37)。野生の植物や園芸植物にこの性質を持つものが多く見られます。雌しべが自己花粉と非自己花粉を識別して、自己花粉の花粉管伸長を特異的に阻止することが分かっていますが、その分子機構の研究は、アブラナを材料とした研究が最も進んでいます。自家不和合性は、S遺伝子座にある花粉側自己認識分子の遺伝子SP11(SCR とも言う)と雌しべ側の受容体の遺伝子SRK によってもたらされ、これら遺伝子は多数の複対立遺伝子を持ちます。2 つの遺伝子間での組換えはほとんど起こらず、SP11 とSRK の1 つの対立遺伝子の組をSハプロタイプと呼びます。花粉と雌しべが同じSハプロタイプのSP11 とSRK によって作られるタンパク質を持っていると、自己花粉と認識され、花粉管伸長が阻害されます。
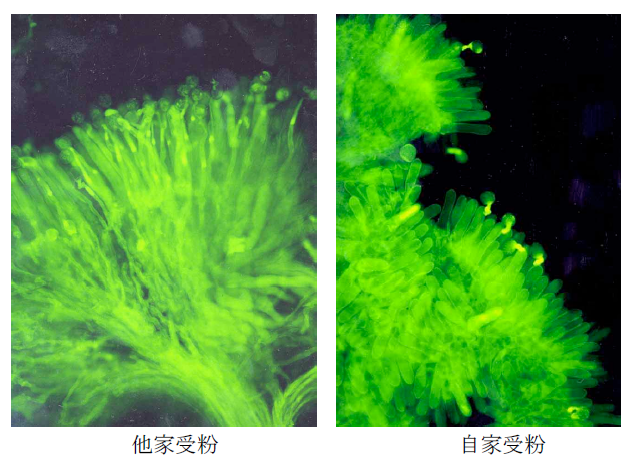
当研究室では、約30 年前にアブラナ属植物の柱頭に極めて多様性が高い糖タンパク質があることを発見し、その糖タンパク質(S糖タンパク質) を単離しました(Nishio and Hinata 1982 Genetics 100, 641-647)。この遺伝子(SLG)の研究から柱頭側の受容体分子の遺伝子SRK が米国で発見されました。SRK 遺伝子の近傍のゲノムの研究から、当研究室で花粉側の自己認識分子の遺伝子SP11 を発見しました(Suzuki et al. 1999 Genetics 153, 391-400)。S遺伝子座の構造はSハプロタイプごとに大きく異なり、SRK とSP11 の距離や遺伝子の向き、遺伝子間領域の塩基配列などが多様です(Fukai et al. 2003 MGG 269, 361-369; Fujimoto et al. 2006 Genetics 173, 1157-1167)。このことが、SRK とSP11 の間での組換えが起こりにくい原因と考えられます。
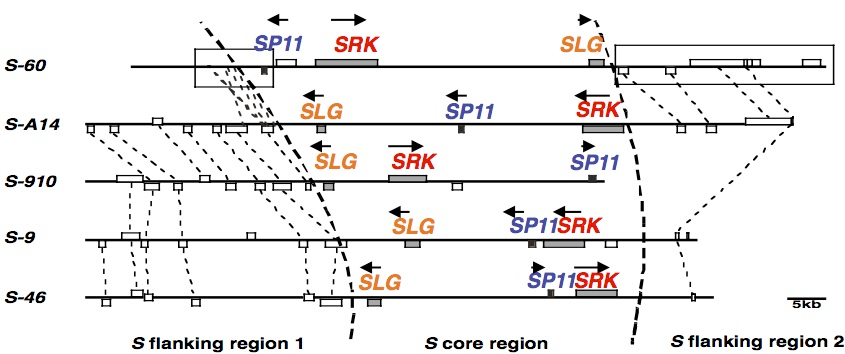
S遺伝子座の構造
ハクサイやカブ(B. rapa)、キャベツやブロッコリー(B. oleracea)、ダイコン(R. sativus)の多数のSハプロタイプのSRK やSP11 の複対立遺伝子の塩基配列を決定し、種が異なっても類似のSハプロタイプが存在することを見出し(Sato et al. 2002 Genetics 162, 931-940)、類似のSハプロタイプは認識特異性も同じであることが分かりました(Kimura et al. 2002 Plant J 29, 215-223)。このことは、小数の個体の突然変異によってこれらの種が生じたのではなく、多数個体の集団が隔離されて異なる種に分化したことを示唆します。異なる種で類似したSハプロタイプを用いることにより、SP11 分子の認識特異性を決定している部位を明らかにしました。また、新しいSハプロタイプが生じる機構のモデルを提唱しました(Sato et al. 2004 Plant Cell 16, 3230-3241)。
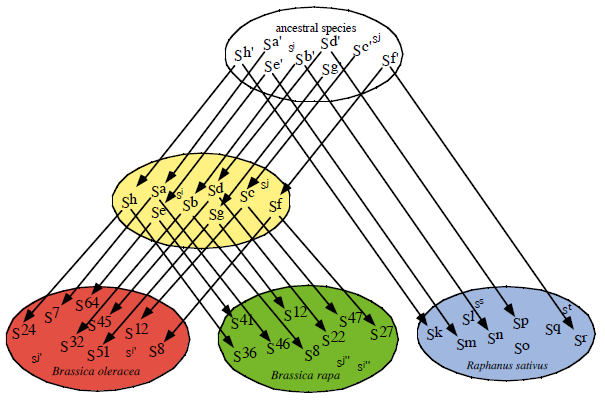
Sハプロタイプは祖先種で既に出来ていたものが現在の種に引き継がれている
アブラナのSハプロタイプの間には優劣関係があります。SP11 の優劣関係は劣性遺伝子側の転写の抑制によることが分かっています。しかし、優性側のSP11 遺伝子が転写されていなくても劣性側SP11 遺伝子の転写抑制が起こることや、優性側のSP11 遺伝子そのものが無くても劣性側SP11 遺伝子の転写抑制が起こること見出しました(Fujimoto et al. 2006 Plant Mol Biol 61, 577-587)。SRK 側の優劣性は転写制御によらないことが分かっていますが、その機構は不明であり、Sハプロタイプの優劣関係はまだまだ謎に満ちています。
U の三角形を構成する複二倍体種が全て自家和合性であり、その祖先ゲノムを単一で持つ種が全て自家和合性であることに着目し、今その原因を調べています。セイヨウナタネ(B. napus)はB. rapa が持つAゲノムとB. oleracea が持つCゲノムを併せ持ちますが、品種によりいくつかのS遺伝子型を持ちます。調べた3 種類のS遺伝子型の自家和合性は、いずれもAゲノム側の優性を示すSハプロタイプの突然変異により生じたことが明らかとなりました(Okamoto et al. 2007 Plant J 50, 391-400; Tochigi et al. 2011 Theor Appl Genet 123, 475-482)。
アブラナでは、1 つの種に約50〜100 種類のSハプロタイプがあると推定されています。B. rapa のSハプロタイプの収集は、約40 年前に当研究室で開始され(Nishio and Hinata 1978 Jpn J Genet 53, 27-33)、その後も継続され(Nou et al. 1993 Sex Plant Reprod 6, 79-86)、現在では約40 種類保有しています。B. oleracea のSハプロタイプは、英国のOckendon 博士より提供を受け、日本の品種に利用されているものも収集して、約50 種類保有しています。ダイコンのSハプロタイプは、約20 種類保有しています。これらは世界で最も充実したSハプロタイプのコレクションであり、B. rapa、B. oleracea、ダイコンのSハプロタイプの増殖は、それぞれカネコ種苗、サカタのタネ、タキイ種苗の協力を得て行いました。
自家不和合性は、アブラナ科野菜の一代雑種品種の採種に利用されていますが、自家不和合性の強度や安定性はSハプロタイプによって異なることから、Sハプロタイプの同定技術が求められています。現在広く利用されているSハプロタイプ同定法は、SLG 遺伝子のPCR−RFLP分析法(Nishio et al. 1996 Theor Appl Genet 92, 388-394)ですが、SLG 遺伝子を持たないSハプロタイプもあることから(Sato et al. 2002)、直接自己認識に関わるSP11 遺伝子の分析による方法を開発しました(Fujimoto and Nishio 2003 Theor Appl Genet 106, 1433-1437)。しかしこの方法はやや高度な分析技術を必要とすることから、改良を加え、より簡易に分析できる方法にしました(Takuno et al. 2010 Theor Appl Genet 120, 1129-1138; Oikawa et al. 2011 Mol Breed 28, 1-12)。
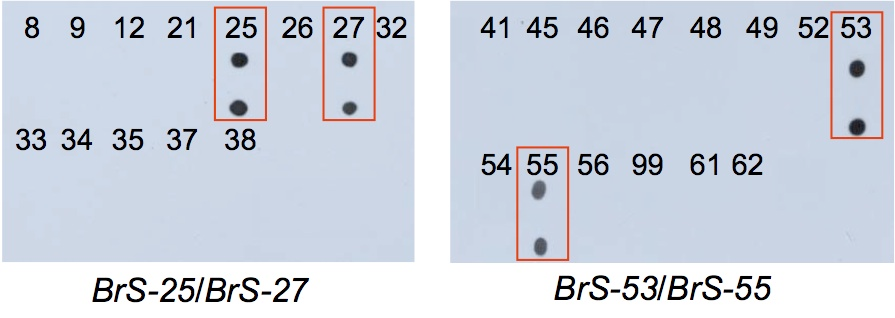
ドットブロット法によるアブラナS遺伝子型の分析例
Sハプロタイプが柱頭と花粉の間で一致すると、自己認識が起こって不和合反応が起こることが分かっていますが、自己認識後どのような機構により不和合反応が起こるかはほとんど分かっていません。当研究室で、B. rapa の自家和合性品種の遺伝分析を行い、S遺伝子座の他にM遺伝子座が自家不和合性に関わることを推定しました(Hinata et al. 1983 Proc Int Rapeseed Conf 1, 354-359)。M遺伝子は自己認識後の機構に関わると考えられます。M遺伝子の実体はMLPK 遺伝子であるとの報告がなされましたが、シロイヌナズナを用いた研究から、この仮説に疑問を示し(Kitashiba et al. 2011 Proc Natl Acad Sci USA 108, 18173-18178)、M遺伝子を探しています。また、チオレドキシンタンパク質がSRK の活性の制御に関わると報告されましたが、この仮説も問題あることを示しました(Yamamoto and Nasrallah 2013 Plant Physiol 163, 1387-1395)。このような自己認識後の分子機構の研究において、米国コーネル大学と協力し、共同研究を続けています。
2. 雄性不稔性の分子機構の研究
細胞質雄性不稔性(Cytoplasmic Male Sterility: CMS)とは雄性器官や雄性配偶体形成時に何らかの欠陥が生じて生殖能力のある花粉ができない性質のことで、母性遺伝する形質です。CMS は自家不和合性と共に一代雑種品種の開発において利用されている非常に重要な性質です。CMS にはミトコンドリアに存在する原因遺伝子と、その原因遺伝子の転写制御や転写後制御をすることで稔性を回復させる核側の稔性回復遺伝子 (Fertility Restorer gene: Rf)が関わっています。特に異質細胞質型のCMS では、由来の異なるミトコンドリアゲノムと核ゲノム間の不和合(障壁)によりCMS が誘発されると考えられています。

D. muralis 細胞質を持つB. oleracea 系統 A:不稔、B:部分不稔、C:可稔
これまで当研究室では、アブラナ科植物の属間での交雑により、細胞質提供親Diplotaxis muralis の細胞質をもつmur 型の異質細胞質雄性不稔性(CMS) B. oleracea を作出しました(日向・今野1979 育雑29, 305-311)。これらを研究材料に、CMS の原因遺伝子を探りました。ミトコンドリアゲノムの解析、遺伝子発現解析の結果、mur 型CMS B. oleracea においてorf72 と名付けたキメラ遺伝子がCMS 原因の有力候補であることを突き止めました(Shinada et al. 2006 Plant Cell Physiol 47, 549-553)。mur 型CMS の稔性回復遺伝子は不明でしたが、多数のDNA マーカーを用いた遺伝分析により、第1 染色体の領域に遺伝子を絞り込み、候補遺伝子を推定しました(Ashutosh et al. 2012 Mol Breed 30, 1781-1792)。
核の遺伝子の突然変異によって雄性不稔となる核遺伝子突然変異は、不稔系統として固定できないことから、一代雑種育種には利用できないと考えられてきました。しかし、雄性不稔性の原因遺伝子を同定し、その突然変異を検出するDNA マーカーを作成しておけば、定植前に不稔個体を選抜でき、一代雑種育種に利用できます。そこでコシヒカリの核遺伝子雄性不稔性突然変異系統の原因遺伝子を調べました。原因遺伝子を第9 染色体の79 kb の中に絞り込み、その中に推定される9 つの遺伝子の塩基配列を調べ、フラボノイド合成に関わる遺伝子の中に1 塩基の欠失(フレームシフト変異)を見出し、それが原因遺伝子であると推定しました(Shirasawa et al. 2013 Mol Breed 31, 805-814)。シロイヌナズナでこの遺伝子の類似遺伝子は、雄性不稔性に関わることが分かっています。この変異を検出するSNP マーカーを作成し、不稔個体を選抜して一代雑種品種を採種するシステムを提案しました。

コシヒカリ雄性不稔性突然変異系統の花と葯(花粉なし)
3. 生殖的隔離機構の研究
生物は種が違う個体の間では、一般に雑種は出来ません。たとえ出来ても、その雑種は子孫を残せません。このような機構を生殖的隔離機構と言います。植物では、異種間で受粉しても受精できない種間不和合性、受精しても雑種種子が出来ない雑種胚退化、雑種種子が出来ても雑種個体が開花まで成長できない雑種致死、開花しても生殖能力のある花粉や胚のうができない雑種不稔が生殖的隔離機構に含まれます。
種間不和合性は、異種間受粉では受精できない機構ですが、アブラナ科では異種の花粉は花粉管が柱頭組織に侵入できず、蕾受粉では受精できるので、自家不和合性と類似しています。B. rapa の種間不和合性が強い品種と弱い品種のF2 を用いて種間不和合性の遺伝分析を行い、種間不和合性強度にS遺伝子座やM遺伝子座は関わっておらず、第2 染色体にこの特性に関わる遺伝子があることを見出しました(Udagawa et al. Theor Appl Genet 121, 689-696)。この原因遺伝子の場所を絞り込み、原因遺伝子を見つけようとしています。
B. rapa とダイコンの属間交雑では、蕾受粉で受精させても雑種胚が退化します。雌親に用いるB. rapa の品種によってはわずかに雑種種子が得られるので、雑種種子が得られる品種と全く得られない品種のF2 を用いて雑種種子形成能の遺伝分析を行い、第1 染色体と第10 染色体に関与遺伝子があることを見出しました。第1 染色体と第10 染色体の関係する領域に、シロイヌナズナで種子形成に関わることが知られているFIE とMSI1 の類似遺伝子があり、それぞれにフレームシフト変異があることを見出しました(Tonosaki et al. 2013 Theor Appl Genet 126, 837-846)。これらの遺伝子の機能の証明をしようとしています。
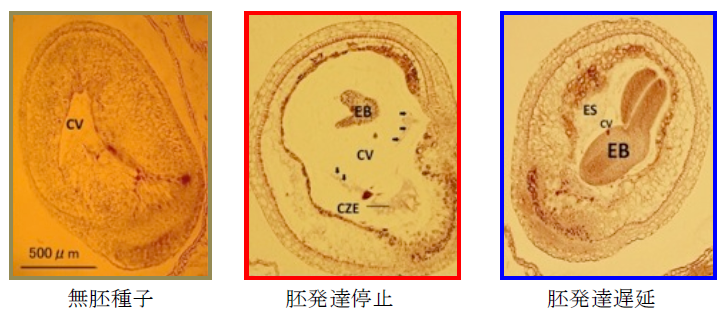
B. rapa とダイコンの属間交雑で見られる胚発達異常
上記以外にも、いろいろやっています。興味のある方は、お気軽にお問い合わせ下さい。お問い合わせ先は、植物遺伝育種学研究室のトップページから。