
Studies on genetics and breeding of environmental stress tolerances
1. Study on cold tolerance gene in rice
Developing a rice cultivar having tolerance to low temperature at booting stage was the most important objective in rice breeding programs in Tohoku region in Japan. Although the global warming proceeds, we still have a cold summer once in several years, and sterile seeds increase by abnormality of pollen development. A rice cultivar ‘Sasanishiki’ was replaced by a cold tolerant cultivar ‘Hitomebore’, but a severe cold summer as that in 1993 may cause yield loss even in ‘Hitomebore’. Further improvement of cold tolerance in rice cultivars is required.
Since ‘Lijiangxintuanheigu’, which is a cultivar in Yunnan province in China, has extremely high cold tolerance at booting stage, a cold-tolerant line ‘Ukei840’, cold tolerance of which is significantly higher that that of ‘Hitomebore’, was developed from a backcross population between ‘Lijiangxintuanheigu’ and ‘Hitomebore’ in the National Agricultural Research Center for Tohoku Region. Collaborating with this center, we analyzed QTLs (quantitative trait loci) for cold tolerance using F2 progeny between ‘Ukei840’ and ‘Hitomebore’ by developing many SNP (single nucleotide polymorphism) markers (Please see “Development of breeding techniques using SNP analysis of genes”.), and detected a QTL at the long-arm end of chromosome 3. Production of many near-isogenic lines having the QTL region of ‘Lijiangxintuanheigu’ with ‘Hitomebore’ background and evaluation of cold tolerance of them enabled delimitation of the genomic region containing a cold tolerance gene into 800 kb. Determining nucleotide sequences of the 800-kb region of ‘Ukei840’ by a next generation sequencer, we detected 150 SNPs between ‘Ukei840’ sequences and the reference genome of ‘Nipponbare’, and selected four candidate genes having important sequence variation (Shirasawa et al. 2012 Theor Appl Genet 124, 937-946). Evaluation of cold tolerance of recombinants between these genes is carried out for identification of the cold tolerance gene. Transgenic rice plants with these candidate genes have already been obtained, but we are unable to perform a reliable cold tolerance test of these plants in an isolated greenhouse special to genetically engineered plants.

Figure. A cold tolerance Chinese cultivar ‘Lijiangxintuanheigu’, toll plants on the left side.
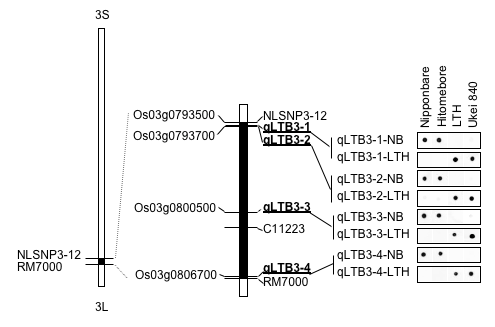
Figure. The chromosomal region harboring a cold tolerance gene of ‘Lijiangxintuanheigu’ and SNP markers in this region.
2. Genetic analysis of high-temperature tolerance at grain-filling period in rice
Extraordinary high temperature in summer of recent years causes deterioration of rice quality, i.e., increase of chalky grains, which became a serious problem for rice production not only in Southwestern region but also in Northeast region (Tohoku region) of Japan. Rice cultivars tolerant to high temperature are required. Although there are differences in the high-temperature tolerance between cultivars, genes responsible for this trait has not been identified. The most sensitive to high temperature evaluated by a frequency of chalky grains was found to be a period from 5 to 10 days after flowering. Populations derived from crossing between lines having the same flowering date have to be used for QTL analysis to identify a high-temperature tolerance gene. ‘Kokoromachi’ and ‘Tohoku 168’ developed in Miyagi Prefectural Furukawa Agricultural Experimental Station, which have the same flowering date, was found to be tolerant and sensitive, respectively, to high temperature. Since recombinant inbred lines have been developed from a cross between ‘Kokoromachi’ and ‘Tohoku 168’ in Miyagi Prefectural Furukawa Agricultural Experimental Station, we collaborated with this experimental station for QTL analysis of high-temperature tolerance. By identifying SNPs between these two lines, many SNP markers were developed. SNP genotyping and evaluation of high-temperature tolerance of each recombinant inbred line enabled identification of one QTL for high-temperature tolerance at a region near the Wx gene on chromosome 6 (Shirasawa et al. 2013 Mol Breed 32, 223-232). However, we have not identified the gene for high-temperature tolerance because of difficulty of evaluation of high-temperature tolerance.

Figure. Extraordinary high temperature in summer causes deterioration of rice quality
3. Drought and salt tolerance studies in Brassica crops
Although we have much rain in Japan, most of countries in the world are suffering for yield loss caused by water deficit due to the global warming and human activities. Many wild plants in Brassica and closely related genera grow in semiarid areas, and are tolerant to drought. Rapeseed (Brassica napus) and mustard (Brassica juncea) are generally tolerant to drought, but drought became a serious problem even for these crops cultivated in China and India. Therefore, supported by Japan Science and Technology Agency, we are collaborating with the Oil Crops Research Institute in Wuhan, China, to develop drought tolerant lines in rapeseed and mustard.
The Great East Japan Earthquake, which occurred on March 11, 2011, inundated more than 20,000 ha of farmland, and excess of salt in the soil caused substantial reduction of crop production. Since Brassicaceae crops are generally tolerant to salt, and we have the Brassicaceae genetic resources, we joined to the Rapeseed Project for Restoring Tsunami-Salt-Damaged Farmland (Please see http://www.nanohana-tohoku.com). Although there was discrepancy in salt tolerance evaluation of rapeseed or Brassicaceae vegetables between previous studies, we found large varietal differences in species of rapeseed and mustard, and selected salt tolerant cultivars or lines. Furthermore, we detected QTLs for salt tolerance by genome-wide association analysis using many rapeseed cultivars and lines, and identified a candidate for the salt tolerance gene.

Rapeseed (Brassica napus) plants growing in a tsunami-salt-damaged field.