2. Pythium oligandrum (PO)施用作物における分子応答機構の解明�
(北海道農業研究センターとの共同研究、平成16年度よりスタート)
【研究の概略】
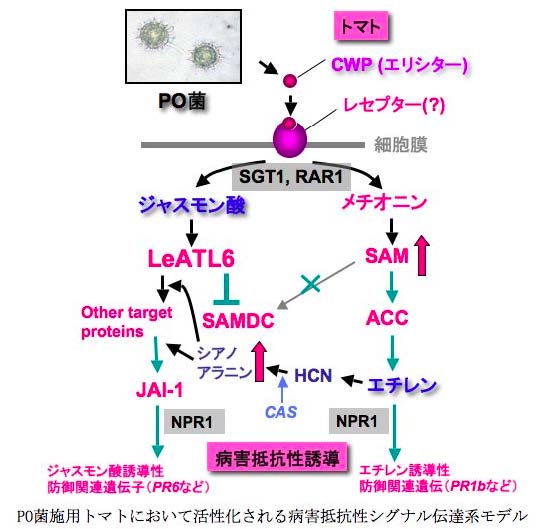
Pythium oligandrum (PO)は, 生物防除微生物として優れた能力を有し, テンサイ苗立枯病, ピーマン半身萎凋病, オオムギBrown rot など多くの土壌病害を抑制する. POによる病害抑制には, POの細胞壁画分に含まれるエリシチン様糖タンパク質(CWP)が重要な役割を果たしていた. CWP 処理トマトにおける青枯病菌に対する抵抗性誘導系をモデルとして, CWP により活性化される防御システムの解析を行なった. cDNA アレイを用いた網羅的な遺伝子発現解析の結果,CWP 処理トマトでは, ジャスモン酸(JA)やエチレン(ET)を介したシグナル伝達系の活性化, 塩基性PRタンパク質遺伝子の発現上昇, シキミ酸経路やファイトアレキシン合成に関与する遺伝子の発現が上昇していた. 一方, JA非感受性変異体トマトでは, CWP によるそれら防御関連遺伝子の発現上昇や, 青枯病菌抵抗性が誘導されなかったことから, JAシグナル伝達系がCWP による抵抗性誘導に必要であると考えられた.
また, CWP 処理により, JA誘導性のユビキチンリガーゼLeATL6 が発現することが見いだされた. このLeATL6 をトマト細胞内で過剰発現させると, 塩基性PR タンパク質遺伝子の発現が誘導されたことから, CWP によるJAシグナルを介した防御システムの活性化には, LeATL6 が重要な役割を果たしていると考えられた. この LeATL6 によりユビキチン化される標的タンパク質を明らかにするため, 酵母two-hybrid system によりLeATL6 と相互作用するタンパク質をスクリーニングしたところ, S-adenosylmethionine decarboxylase (SAMDC)が単離された. 実際に、CWP 処理トマトでは, SAMDC 活性の低下が認められるとともに, ET生成量が一過的に上昇していた. SAMDC の基質であるS-adenosylmethionine は, ET の生合成の前駆物質でもあることから, JA 誘導性LeATL6 を介したSAMDC 活性の低下と, ET シグナル伝達系の活性化との関連に興味が持たれた.
さらに, 防御応答に異常をきたしたシロイヌナズナ変異体を用いた解析から, CWP による防御関連遺伝子の発現や細菌病抵抗性の誘導は, 全身獲得抵抗性(SAR)やRhizobacteria
による誘導抵抗性(ISR)の発現に関わるNPR1 遺伝子, Rタンパク質による病害抵抗性に関わるSGT1, RAR1 遺伝子などに依存していた. 以上より, P. oligandrum により誘導される防御システムでは, それらの遺伝子に依存したJA/ETシグナル伝達系の活性化により制御されていることが明らかになった.
【原著論文リスト】(2005-2012 年)
Ishihara, T., Mitsuhara, I., Takahashi, H. and Nakaho, K. (2012) Transcriptome
analysis in resistant and susceptible cultivars of tomato infected with
Ralstonia solanacearum. PLoS One (in press)
Ikeda, S., Shimizu, A., Shimizu, M., Takahashi, H. and Takenaka, S. (2012) Biocontrol of black scurf on potato by seed tuber treatment with Pythium oligandrum. Biological Control 60: 297–304
Takenaka, S., Yamaguchi, K., Masunaka, A., Hase, S., Inoue, T. and Takahashi, H. (2011) Implications of oligomeric forms of POD-1 and POD-2 proteins isolated from cell walls of the biocontrol agent Pythium oligandrum in relation to their ability to induce defense reactions in tomato. Journal of Plant Physiology 168: 1972-1979.
Masunaka, A., Sekiguchi, H., Takahashi, H. and Takenaka, S. (2010) Distribution and expression of elicitin-like protein genes of the biocontrol agent Pythium oligandrum. Journal of Phytopathology 158: 417-426.
Takahashi, H., Hondo, D., Hase, S. and Takenaka, S. (2010) Identification of a protein
that interacts with LeATL6 ubiquitin-protein ligase E3 upregulated in tomato treated with elicitin-like cell wall proteins of Pythium oligandrum. Journal of Phytopathology 158: 132-136.
Kawamura, Y., Takenaka, S., Hase, S., Kubota, M., Ichinose, Y., Kanayama,
Y., Nakaho, K. and Takahashi, H. (2009) Enhanced defense responses in Arabidopsis induced by the cell wall protein fractions from Pythium oligandrum require SGT1, RAR1, NPR1 and JAR1. Plant Cell Physiology 50: 924-934.
Kawamura, Y., Hase, S., Takenaka, S., Kanayama, Y., Yoshioka, H., Kamoun,
S. and Takahashi, H. (2009) INF1 Elicitin activates jasmonic acid-and ethylene-mediated signalling pathways and induces resistance to bacterial wilt disease in
tomato. Journal of Phytopathology 157: 287-297.
Masunaka, A., Nakaho, K., Sakai, M., Takahashi, H. and Takenaka, S. (2009)
Visualization of Ralstonia solanacearum cells during biocontrol of bacterial wilt disease in tomato with Pythium oligandrum. Journal of General Plant Pathology 75: 281-287.
Hase, S., Takahashi, S., Takenaka, S, Nakaho, K., Arie, T., Seo, S., Ohashi, Y. and Takahashi, H. Jasmonic acid signaling is required for bacterial wild disease resistance induced by biocontrol agent Pythium oligandrum in tomato. Plant Pathology 57:870?876 (2008).
Takenaka, S., Sekiguchi, H., Nakaho, K., Tojo, M., Masunaka, A. and Takahashi,
H. (2008) Colonization of Pythium oligandrum in the tomato rhizosphere for biocontrol of bacterial wilt disease analyzed by real-time PCR and confocal laser scanning microscopy. Phytopathology 98:187-195.
Hondo, D., Hase, S., Kanayama, Y., Yoshikawa, N., Takenaka, S. and Takahashi, H. (2007) Up-regulation of LeATL6 that encodes a fungal elicitor-responsive ubiquitin ligase induces jasmonic acid-dependent proteinase inhibitor gene expression in tomato. Molecular Plant-Microbe Interactions 20: 72-81.
Hase, S., Shimizu, A., Nakaho, K., Takenaka, S. and Takahashi, H. (2006) Induction of transient ethylene and reduction in severity of tomato bacterial wilt by
Pythium oligandrum. Plant Pathology 55: 537-543.
Takahashi, H., Ishihara, T., Hase, S., Chiba, A., Nakaho, K., Arie, T.,
Teraoka, T., Iwata, M., Tugane, T., Shibata, D. and Takenaka, S. (2006) Beta-cyanoalanine synthase as a molecular marker induced resistance by fungal glycoprotein elicitor and
commercial plant activators. Phytopatjhology 96: 908-916.
Takenaka, S., Nakamura, Y., Ono, T., Sekiguchi, H., Masunaka, A. and Takahashi, H. (2006) Novel elicitin-like proteins isolated from the cell wall of the biocontrol agent Pythium oligandrum induce defence-related genes in sugar beet. Molecular Plant Pathology 7: 325-339.
Takahashi, H., Shimizu, A., Arie, T., Rosmalawati, S., Fukushima, S., Kikuchi, M., Hikichi, Y., Kanda, A., Ohnishi, K., Ichinose, Y., Yasuda, C., Kodama, M.,
Egusa, M., Masuta, C., Sawada, H., Shibata, D., Hori, K. and Watanabe, Y. (2005)
Catalog of Micro-Tom tomato responses to common fungal, bacterial and viral pathogens. Journal of General Plant Pathology 71: 8-22.
【総説・著書リスト】(2009-2010年)
竹中重仁・出崎里永子・高橋英樹(2010)「非病原菌Pythium oligandrum を利用した広スペクトル微生物農薬の開発」バイオインダストリー27: 39-46.
Takahashi, H. and Takenaka, S. (2010) “Defense system induced by elicitin-like proteins of biocontrol agent Pythium oligandrum”. Genome-enabled integration of research in plant-pathogen systems. In 10th Japan-USA Scientific Seminar (Eds. T. Walpert, T. Shiraishim, K. Akimitsu and J. Glazebrook), APS Press, St Paul,
Minnesota, USA (in press).
高橋英樹(2010)「抵抗性発現に関わる遺伝子制御」植物病理学(眞山滋志・難波成任編)pp224-228, 文英堂高橋英樹・長谷修・竹中重仁(2009)「モデル植物トマト:マイクロトムを用いた病害研究の現状と今後の展望」植物防疫63, 605-607.
高橋英樹・竹中重仁(2009)「土壌生息菌Pythium oligandrum が生産するエリシチン様タンパク質により誘導される感染防御システムの分子機構」日本農薬学会誌34, 39-341
高橋英樹・竹中重仁(2009)「遺伝子発現からみた誘導抵抗の全体像」微生物と植物の相互作用-病害と生物防除-(百町満朗・対馬誠也編)第2章2-1,
pp128-133, ソフトサイエンス社
研究の詳細
【研究の背景】Pythium oligandrum (PO)は卵菌類に属する非病原性の土壌生息菌であり、テンサイ苗立枯病、ピーマン半身萎凋病、オオムギbrown rot など多くの土壌病害に対して抑制効果がある。POは菌寄生性や、根圏域で栄養と生息域を占める能力が高い特性を有することから、病害抑制は主として病原菌に対する直接的な拮抗作用によるものと考えられてきた。その後、トマト、テンサイ、コムギを用いた実験から、POはエリシター分子を介して病害抵抗性を誘導する能力も有していることが明らかとなった。POの細胞壁分画からは、テンサイとコムギに対して抵抗性誘導能を活性化させる細胞壁タンパク質画分(CWP) が精製されている。CWP は2種類の主要タンパク質(POD-1とPOD-2)からなり、両タンパク質とも約15%の糖(GlcNAc, GalNAc, Glu) を含む糖タンパク質である(Takenaka et al., 2006)。また、POD-1とPOD-2はアミノ酸レベルで83%の相同性があり、中央部に3つのElicitin signature を含むエリシチンドメインとC末にO-グリコシド結合型のドメインが存在した。この特徴からPOD-1とPOD-2は新しいタイプのエリシチン様タンパク質であることが明らかとなった。CWP はテンサイとコムギの他にもナス科のトマト、ジャガイモ、モデル植物であるシロイヌナズナ、イネなどの多くの植物に抵抗性誘導活性を示すことが報告されている(Takahashi et al., 2006; Hase et al., 2006, 2008; Kawamura et al., 2009)。
【研究の目的】
トマトのモデル植物であるマイクロトム(Takahashi et al., 2005)を用いて、PO菌施用あるいは細胞壁タンパク質エリシター (CWP)の処理による植物の遺伝子発現の変動と病害抑制に関わる植物のシグナル伝達系の解析を行い、抵抗性誘導の分子機構を解明する。
�
【研究成果】
(1) PO 菌施用トマトにおける遺伝子発現変動の解析
① PO 菌細胞壁タンパク質(CWP)エリシターを処理したトマト根における遺伝子発現変動の網羅的解析:PO施用による誘導抵抗性の分子機構を解析するため、POが生産する細胞壁タンパク質エリシター (CWP)処理に対して発現が変動する宿主遺伝子の解析を行った。マイクロトムの根部にCWP および蒸留水(DW)を処理し、4時間後に抽出したRNA から調製したプローブを用いて、マイクロトムEST 約11,000 クローンをスポットしたマクロアレイにより遺伝子発現解析を行った(Takahashi et al., 2006) 。その結果、DW処理に対してCWP 処理において142遺伝子で発現が3倍以上増加し、99遺伝子で発現が1/3 以下に減少していた。また、それら遺伝子の中には、ジャスモン酸(JA)やエチレンを介したシグナル伝達系に関与すると考えられる遺伝子、糖代謝やアミノ酸代謝に関わる遺伝子、ファイトアレキシン合成などの2次代謝系に関わる遺伝子などが含まれていた(表1) 。さらに、PO菌を施用したトマト根における遺伝子発現変動を同様にアレイ解析したところ、発現が3倍以上に上昇したものが68遺伝子、1/3 以下に減少したものが73遺伝子認められた。また、その結果と、CWP エリシター処理における遺伝子発現変動の結果を比較したところ、両処理の間で共通して発現が変動する遺伝子が多数見出された。したがって、PO菌処理とCWP エリシター処理においては、共通した分子応答機構が活性化されていることが確認された。

② PO菌あるいはCWP エリシター処理イネおよびジャガイモにおける遺伝子発現変動解析: PO菌はトマト青枯病以外にイネ苗立枯病、イネ籾枯細菌病、ジャガイモ黒あざ病に高い防除効果を示すことが明らかになっている。防除効果が認められた作物の間で、PO菌により発現が変動する遺伝子を網羅的に比較解析すれば、誘導抵抗性に関わる遺伝子を絞り込むことができると考え、CWP 処理イネにおける遺伝子発現の網羅的解析、および得られたデータをCWP処理トマトにおけるデータと比較解析した(表1)。その結果、トマトとイネに共通して、塩基性PRタンパク質遺伝子(basic PR-1、basic chitinase など)、ファイトアレキシン合成に関わる2次代謝系遺伝子[イネのファイトアレキシン合成に関わるferulate-5-hydroxylase (FAH1) やkaurene synthase (KS)、トマトのファイトアレキシン合成に関わるfarnesyl pyrophosphate synthase 、shikimate 5-dehydrogenase 、chorismate synthase ]、ジャスモン酸合成に関わるalleneoxide synthase (AOS)やlipoxygenase (LOX)、RING-H2 finger 型ユビキチンリガーゼの発現が顕著に上昇することが明らかになった(表1)。一方、全身獲得抵抗性(SAR)において重要な役割を果たすとされるサリチル酸シグナル伝達系によって制御されている酸性PRタンパク質遺伝子やサリチル酸の生合成に関わる遺伝子については、CWP 処理トマト、イネいずれにおいても明確な発現変動は認められなかった。また、ジャガイモについては、塊茎ディスクにCWP エリシターを処理し、トマトにおいて発現が上昇しているDAHP 、LOX 、PR-6遺伝子についてノザン法により発現を解析したところ、いずれもCWP エリシター処理により顕著に発現が上昇した。したがって、PO菌施用トマト、イネ、ジャガイモでは、PO菌による病害抵抗性の誘導において、CWP エリシターを介してジャスモン酸・エチレンシグナル伝達系の活性化と塩基性PRタンパク質遺伝子やファイトアレキシン合成系の遺伝子発現が誘導されることが明らかとなった。
(2) PO 施用トマトにおいて病原体接種後の抵抗性誘導過程で発現変動する遺伝子の解析
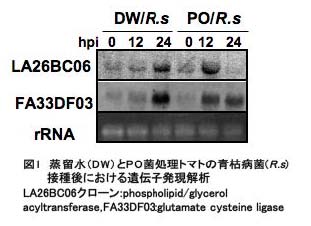
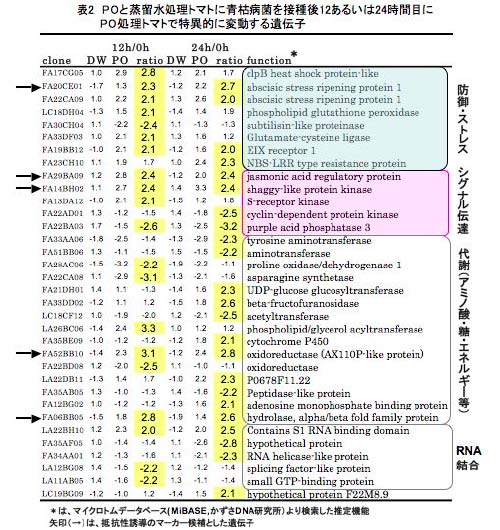
本研究では、PO菌を施用したマイクロトムに青枯病菌を接種し、抵抗性誘導過程における宿主遺伝子発現変動を、12,158 のマイクロトムEST クローンがスポットされたcDNA マクロアレイを用いて解析した。PO菌糸破砕液と蒸留水(DW)を処理した5日目のトマトの根部に青枯病菌を接種し、0, 12および24時間目における発現を解析した結果、0時間目に比べ12あるいは24時間後にPO処理トマトでのみ特異的に2倍以上増加ないし減少した遺伝子は59存在した。これらの遺伝子の発現が、DWを処理した対照とPO処理トマトの間でどの程度の差があるかを調べるために、一部のクローンについてノザン解析を行った。図1は、POおよびDW処理トマトにおける、クローンLA26BC06 (Glutamate cysteine ligarse)とFA33DF03 (phospholipid/glycerol acyltransferase)のノザン解析結果である。両遺伝子の発現は、青枯病菌接種後12時間目で、DW処理した対照に比べてPO処理トマトで高いことが確かめられた(図1)。発現変動が認められた59遺伝子の中で、25遺伝子は機能未知タンパク質をコードしており、34遺伝子は、防御遺伝子を含むストレス応答、シグナル伝達、アミノ酸や糖およびエネルギー代謝に関与するものと推定された(表2) 。この中にはEthylene induced xylanase (EIX)のレセプター遺伝子ホモログやjasmonic acid regulatory protein 遺伝子ホモログが含まれており、PO菌をあらかじめ処理したトマトは、青枯病菌の接種後もエチレンとジャスモン酸シグナル伝達系の調節を受けていることが示唆された。
(3) PO菌施用により活性化されるシグナル伝達系と発現上昇する遺伝子産物の機能解析
①PO菌およびPO菌細胞壁タンパク質(CWP)エリシター処理トマトにおけるサリチル酸、ジャスモン酸、エチレンを介したシグナル伝達系の活性化
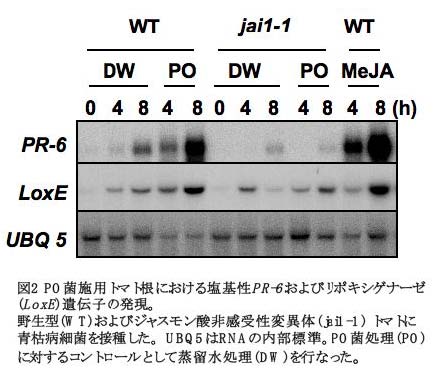
・ジャスモン酸シグナル伝達系: PO菌をトマト根に処理し、4および8時間後におけるジャス モン酸(JA) 誘導性遺 伝子(塩基性PR-6およ びLoxE)の発現を解析 したところ、いずれ も顕著な発現上昇が 認められた( 図2) (Hase et al., 2008)。ま た、PO菌施用による これらの遺伝子の発 現は、JA非感受性変 異体トマトである jai1-1では認められな かった(図2)。さらに、 PO 菌を施用した jai1-1変異体では、コ ントロールと比較し て、青枯病菌に対する抵抗 性がほとんど誘導されな かった(図3)。したがって、 JAを介したシグナル伝達 系は、PO菌施用により誘 導される青枯病菌抵抗性 の発現において重要な役 割を果たしているものと 考えられた。

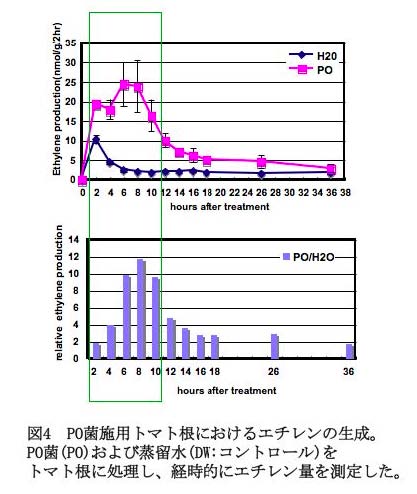
- ・エチレンシグナル伝達系:PO菌をマイクロトムの根に処理し、エチレン生成量を経時的に測定したところ、対照の蒸留水処理に比べて、処理後数時間で生成量に有意な増加が認められた(図4) (Hase et al., 2006)。また、エチレン誘導性PR遺伝子群(塩基性PR1,PR3)の発現をノザン法により解析した結果、いずれもPO菌を処理した根では、対照に比べて高い発現を示した。さらに、本菌の細胞壁タンパク質エリシター (CWP) を根に処理した場合も、エチレン生成量の有意な増加、ならびにPR遺伝子発現の増加が認められたことから、PO菌を施用したトマト根では、JAシグナル伝達系に加え、エチレンを介したシグナル伝達系も活性化されている可能性が考えられた。 ・サリチル酸シグナル伝達系: PO菌をトマト根に処理後4および8時間後におけるサリチル酸(SA)量を測定したところコントロールとの間に優位な差は認められなかった。また、SA誘導PR遺伝子であるPR-2aの発現をノザン解析したが、処理後4および8時間後において遺伝子の発現は認められなかった(Hase et al., 2008)。さらに、SA水酸化酵素遺伝子(nahG)を過剰発現さることにより細胞内におけるSAの蓄積を常に抑制した形質転換トマトにおいても、PO処理により青枯病菌に対する誘導抵抗性は発現した。これらの結果から、PO処理によってSAシグナル伝達系は活性化されず、青枯病菌に対する抵抗性誘導にSAは必要でない可能性が高いと判断された。以上のように、PO菌処理トマト根において、3種類のシグナル物質を介したシグナル伝達系の活性化について解析した結果、PO菌処理による誘導抵抗性発現には、JA
およびエチレンを介したシグナル伝達系が関与していることが明らかになった。
- ② 発現が上昇する遺伝子産物の機能解析・RING-H2 zinc-finger protein 遺伝子: PO菌施用トマトとCWP エリシター処理トマトおよびイネにおける遺伝子発現解析においてRING-H2 zinc-finger モチーフをもつタンパク質(LeATL6)
をコードする遺伝子の発現が上昇していることが見出された(表1)。RING-H2 zinc-finger モチーフは、標的タンパク質の特異的分解に関わるユビキチンリガーゼに認められるモチーフであることが最近明らかになってきている。RING-H2
zinc-finger 型ユビキチンリガーゼは、シロイヌナズナゲノム上に約300 コピー存在し、形態形成や環境応答など様々な生理現象において重要な役割を果たしているものと考えられている。そこで、PO
菌およびCWP エリシター処理により発現が顕著に上昇するLeATL6 の機能について解析を行なった。
 LeATL6 のcDNA をクローニングし、データベースにより解析を行なったところ、トマトのLeATL6 は、キチンエリシターで発現が誘導されるシロイヌナズナATL6 ユビキチンリガーゼと高い相同性と有することが明らかになった。次に、LeATL6 のユビキチンリガーゼ活性を確認するため、大腸菌で発現させたLeATL6
タンパク質を用いてin vitro で活性を測定したところ、ポリユビキチン化活性を有することが明らかになった。シグナル伝達系変異体およびcDNA アレイを用いた遺伝子発現の網羅的解析から、ジャスモン酸(JA)
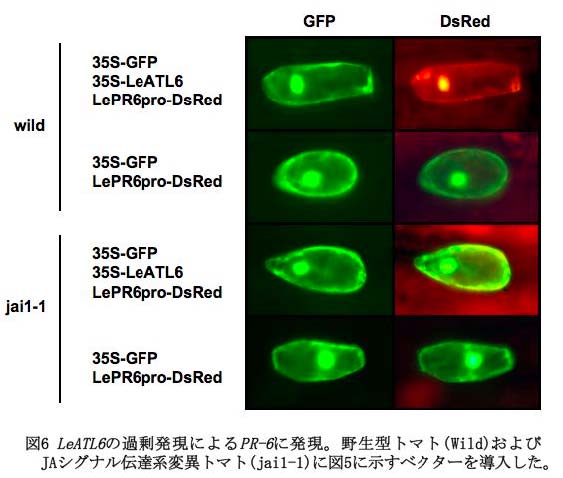
を介したシグナル伝達系がPO菌施用による誘導抵抗性において主要な役割を果たしているものと考えられる。 そこで、一過的遺伝子発現システムを用いて、LeATL6 の発現上昇とJAシグナル伝達系の活性化の関係を調べた。図5に示すように、 JAシグナル伝達系により発現が制御されている防御関連タンパク質遺伝子のひとつである塩基性PR-6遺伝子のプロモーター配列にレポーターとしてDeRed コード領域とポリA付加配列(NOST)を連結したベクター(PR-6pro:DsRed:NOST)を構築した。そのベクターをCaMV35S プロモーターにLeATL6 を連結したベクター(CaMV35Spro:LeATL6:NOST)とともに、パーティクルガン法でトマト葉表皮細胞に導入した。遺伝子導入細胞は、CaMV35S プロモーター:GFP:NOST ベクター(CaMV35Spro:GFP:NOST)をさらに同時に導入することにより、緑色蛍光で判定した。ベクター導入細胞では、CaMV35S プロモーターによりLeATL6 が一過的に過剰発現されるが、LeATL6 の発現によりJAシグナル応答遺伝子であるPR-6の発現が誘導されるならば、細胞はレポーターであるDsRed の発現によって赤色蛍光を発することになる( 図5参照)。実験の結果、PR-6pro:DsRed:NOST とCaMV35Spro:LeATL6:NOST を同時に導入した場合は、PR-6pro:DsRed:NOST を単独で導入した場合と比較して、赤色蛍光を発する細胞が有意に増加していた(図6)(Hondo et al., 2007) 。さらに、同様の実験を、JAシグナル伝達系変異体であるjai1-1で行なったところ、PR-6pro:DsRed:NOST とCaMV35Spro:LeATL6:NOST を同時に導入した場合とPR-6pro:DsRed:NOST を単独で導入した場合で、赤色蛍光を発する細胞数に差は認められなかった(図6)。以上の結果より、LeATL6 の過剰発現は、JA シグナル伝達系を介して防御関連遺伝子であるPR-6の発現を制御しているものと考えられた。
LeATL6 のcDNA をクローニングし、データベースにより解析を行なったところ、トマトのLeATL6 は、キチンエリシターで発現が誘導されるシロイヌナズナATL6 ユビキチンリガーゼと高い相同性と有することが明らかになった。次に、LeATL6 のユビキチンリガーゼ活性を確認するため、大腸菌で発現させたLeATL6
タンパク質を用いてin vitro で活性を測定したところ、ポリユビキチン化活性を有することが明らかになった。シグナル伝達系変異体およびcDNA アレイを用いた遺伝子発現の網羅的解析から、ジャスモン酸(JA)
を介したシグナル伝達系がPO菌施用による誘導抵抗性において主要な役割を果たしているものと考えられる。 そこで、一過的遺伝子発現システムを用いて、LeATL6 の発現上昇とJAシグナル伝達系の活性化の関係を調べた。図5に示すように、 JAシグナル伝達系により発現が制御されている防御関連タンパク質遺伝子のひとつである塩基性PR-6遺伝子のプロモーター配列にレポーターとしてDeRed コード領域とポリA付加配列(NOST)を連結したベクター(PR-6pro:DsRed:NOST)を構築した。そのベクターをCaMV35S プロモーターにLeATL6 を連結したベクター(CaMV35Spro:LeATL6:NOST)とともに、パーティクルガン法でトマト葉表皮細胞に導入した。遺伝子導入細胞は、CaMV35S プロモーター:GFP:NOST ベクター(CaMV35Spro:GFP:NOST)をさらに同時に導入することにより、緑色蛍光で判定した。ベクター導入細胞では、CaMV35S プロモーターによりLeATL6 が一過的に過剰発現されるが、LeATL6 の発現によりJAシグナル応答遺伝子であるPR-6の発現が誘導されるならば、細胞はレポーターであるDsRed の発現によって赤色蛍光を発することになる( 図5参照)。実験の結果、PR-6pro:DsRed:NOST とCaMV35Spro:LeATL6:NOST を同時に導入した場合は、PR-6pro:DsRed:NOST を単独で導入した場合と比較して、赤色蛍光を発する細胞が有意に増加していた(図6)(Hondo et al., 2007) 。さらに、同様の実験を、JAシグナル伝達系変異体であるjai1-1で行なったところ、PR-6pro:DsRed:NOST とCaMV35Spro:LeATL6:NOST を同時に導入した場合とPR-6pro:DsRed:NOST を単独で導入した場合で、赤色蛍光を発する細胞数に差は認められなかった(図6)。以上の結果より、LeATL6 の過剰発現は、JA シグナル伝達系を介して防御関連遺伝子であるPR-6の発現を制御しているものと考えられた。
PO菌施用による抵抗性誘導におけるユビキチンリガーゼLeATL6 の役割を明らかにするためには、LeATL6 の標的タンパク質の同定が必要である。LeATL6 によりユビキチン化されるトマトの標的タンパク質を明らかにするため、酵母two-hybrid 系によりLeATL6 と相互作用するタンパク質の同定を試みた。約1x106クローンについてスクリーニングを行なったところ、最終的にS-adenosylmethionine decarboxylase (SAMDC)が単離された(図7)(Takahashi et al., 2009 )。SAMDC は、LeATL6 が標的タンパク質と結合すると推定されるC末端可変領域とも相互作用することが確認された。ヒトのSAMDC は、26Sプロテアソームにより分解されるとの報告がすでにあることからも、細胞内のSAMDC 量が、ユビキチンープロテアソーム系により調節されていることが示唆された。SAMDC がLeATL6 によりユビキチン化され、プロテアソーム系により分解されるとすれば、CWP 処理トマトにおいてはSAMDC 活性が低下することが予想される。実際に、SAMDC 活性の変動を直接測定したところ、CWP 処理トマト根においてSAMDC 活性が有意に低下することが明らかになった(図8)。SAMDC は、エチレンの前駆体でもあるS-アデノシルメチオニン(SAM)からスペルミン、スペスミジン合成に関わることから、LeATL6 の標的タンパク質であるSAMDC の活性の低下が、SAM の相対量を増加させることによりエチレンシグナル伝達系の活性化に関与しているものと考えられた。SAMDC は、エチレンの前駆体でもあるS-アデノシルメチオニン(SAM)からスペルミン、スペスミジン合成に関わることから、LeATL6 の標的タンパク質であるSAMDC の活性の低下が、SAM の相対量を増加させることによりエチレンシグナル伝達系の活性化に関与しているものと考えられた。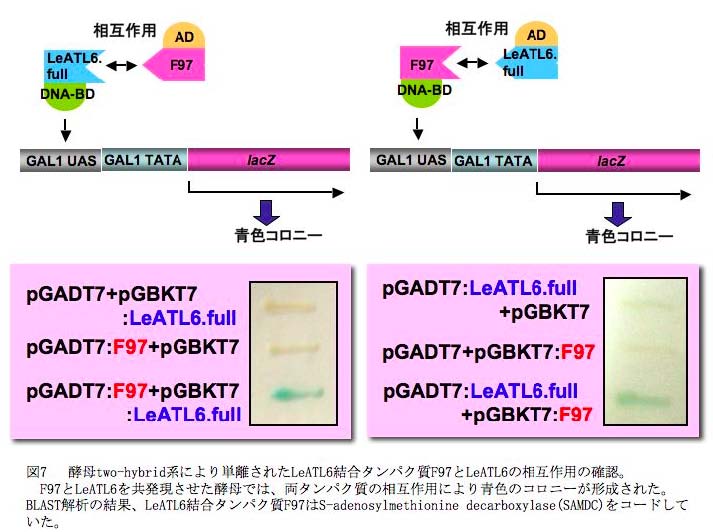
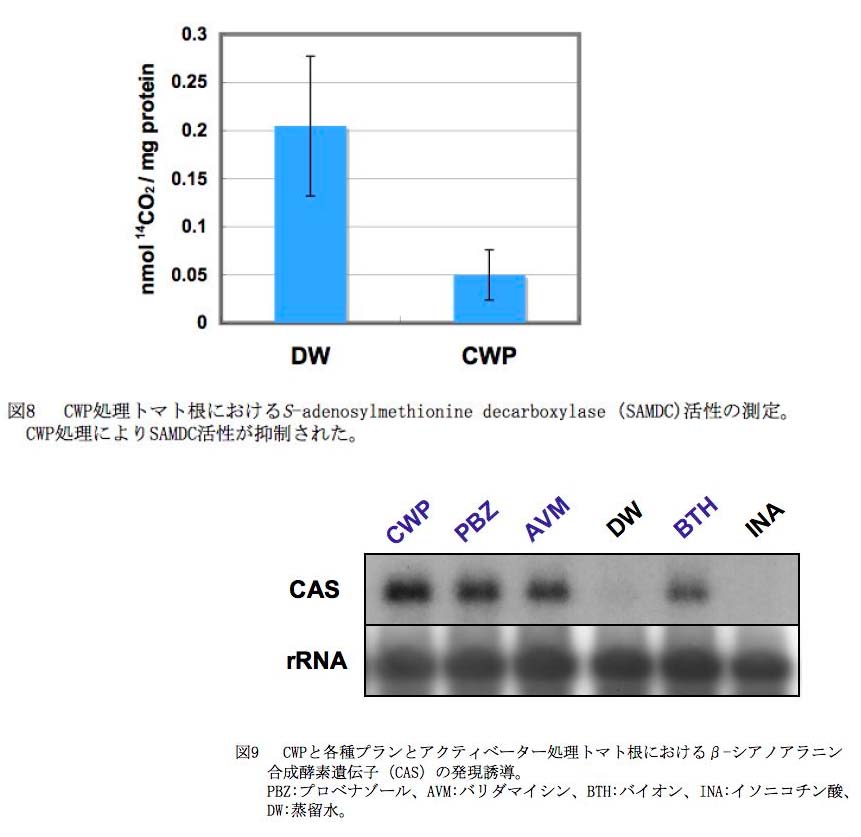
・Beta-cyanoalanine synthase 遺伝子: CWP 処理トマト根において発現が変動する遺伝子の中から、特にコントロールと比較して7倍以上の顕著な発現上昇が認められた9遺伝子を選び、さらに解析を進め、安定して発現が上昇するβ-シアノアラニンシンターゼ(CAS)遺伝子を選抜した(Takahashi et al., 2006)。このCAS 遺伝子は、(i)病害抵抗性の誘導剤として実用化されている3 種類のPlant activator (プロベナゾールやバイオン)の処理に対しても顕著な発現上昇を示し( 図9)、一方(ii)他の環境ストレスでは誘導されなかった。また、 (iii)PO 菌(卵胞子+ 菌糸破砕物)、北海三共液剤(Lot503251) 、北海三共液粉剤(Lot50427)の処理によってもCAS の発現が誘導された。さらに、(iv)CWP 処理によるCAS 遺伝子の発現上昇の経時変動、(vi)CWP 処理によるCAS 遺伝子の組織特異性などについて検討を行い、CAS 遺伝子が本コンソーシアムにおけるPO製剤の開発において、病害抵抗性誘導のマーカー遺伝子として利用できることを明らかにした。また、PO菌施用による抵抗性誘導におけるCAS の役割について解析を行った。CAS は、PO菌による抵抗性誘導におけるエチレン生成の上昇において、その生成副産物であるシアンを解毒化する役割を果たしていると推察される。このCAS によるシアン解毒により、生成するシアノアラニンが、抵抗性誘導において機能しているかどうかを解析するため、シアノアラニンをトマト根に処理して遺伝子発現を解析した。その結果、シアノアラニン処理により、サリチル酸応答性の防御関連遺伝子は発現せず、ジャスモン酸応答性の防御関連遺伝子(PR-6など)の発現が顕著に誘導されることが明らかになった(図10) 。したがって、PO菌施用トマトでのエチレン生成に伴うシアノアラニンの上昇が、ジャスモン酸シグナル伝達系の活性化に関与している可能性が考えられた。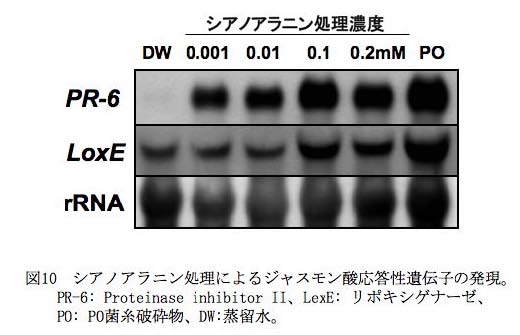
-
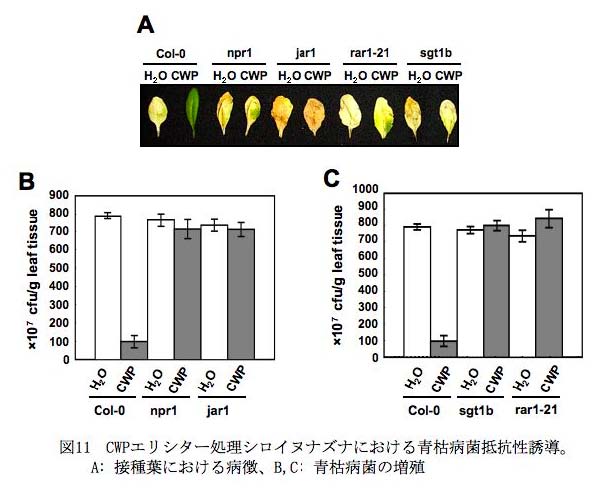
- (4) 防御応答に異常をきたしたシグナル伝達系変異体を用いたPO菌による抵抗性誘導機構の解析 : トマトの遺伝子はゲノムあたり30,000以上あると見積られているが、その70%の遺伝子はシロイヌナズナの遺伝子と共通の機能を有していると推定されている。CWP によるトマトの抵抗性誘導機構を詳細に解明していくためには、変異体が整備されているシロイヌナズナを用いた研究が重要であることから、CWP を処理したシロイヌナズナにおける遺伝子発現を解析した。その結果、PO菌またはCWP エリシター処理シロイヌナズナではJA誘導性の防御遺伝子(PDF1.2)が誘導され、JA非感受性変異体jar1 とcoi1 では誘導されなかったことから、シロイヌナズナではトマトと同様に、CWP エリシター処理によりJA のシグナル伝達系が活性化されると考えられた(Kawamura
et al., 2009) 。さらに抵抗性シグナル伝達系に異常をきたした12種類の変異体を用いた解析から、NB(nucleotide-binding)-LRR(leucine-rich
repeat) ドメインをもつRタンパク質を介する抵抗性に関与するsgt1 とrar1 変異体、およびSARやリゾバクテリアによる誘導抵抗性(ISR)に異常をきたした変異体npr1 でも、CWP によるPDF1.2 遺伝子の発現が誘導されないことが明らかになった。さらに、CWP エリシターを処理した野生型シロイヌナズナではPseudomonas syringae pv. tomato DC3000 および青枯病菌に対する抵抗性が誘導されたが、jar1 、coi1 、sgt1 、rar1 、およびnpr1 変異体では同抵抗性が失われた(図11)(Kawamura et al., 2009 )。これらの結果から、SGT1 とRAR1 ならびにNPR1 が、PO菌ならびにCWP エリシターによる抵抗性誘導にも関与していると考えられた。以上の結果を総合して、研究の概略図にPO菌による抵抗性誘導の分子機構のモデルを示した。